Back to Don's Maps
 La Grotte de la Vache is important for the complete camp of Magdalenian hunters found, and may be seen almost as it was 12 000 to 15 000 years ago. It was the living area for the artists of Niaux, just across the valley. Weapons, tools, typical game and artworks have been recovered from this small but important site.
La Grotte de la Vache is important for the complete camp of Magdalenian hunters found, and may be seen almost as it was 12 000 to 15 000 years ago. It was the living area for the artists of Niaux, just across the valley. Weapons, tools, typical game and artworks have been recovered from this small but important site.  Niaux Cave, or la Grotte de Niaux is one of the most famous prehistoric caves in Europe. It lies in the northern foothills of the Pyrenees, and is located in Ariège, in the valley of Vicdessos, across the valley from the smaller Grotte de la Vache where the artists lived over the winter ibex season, in an area rich with prehistoric sites. The huge cave entrance, 55 metres high and 50 metres wide, is at 678 metres above sea level. There are more than two kilometres of galleries, with a hundred or more superb paintings from Magdalenian times, most of which are in the famous 'Salon Noir', 800 metres from the entrance. Many of the paintings are done in the classic style of the Magdalenian, outlined in black or red pigment, mostly haematite or manganese dioxide respectively.
Niaux Cave, or la Grotte de Niaux is one of the most famous prehistoric caves in Europe. It lies in the northern foothills of the Pyrenees, and is located in Ariège, in the valley of Vicdessos, across the valley from the smaller Grotte de la Vache where the artists lived over the winter ibex season, in an area rich with prehistoric sites. The huge cave entrance, 55 metres high and 50 metres wide, is at 678 metres above sea level. There are more than two kilometres of galleries, with a hundred or more superb paintings from Magdalenian times, most of which are in the famous 'Salon Noir', 800 metres from the entrance. Many of the paintings are done in the classic style of the Magdalenian, outlined in black or red pigment, mostly haematite or manganese dioxide respectively.Grotte de la Vache near Niaux - A study of the fauna and occupation by humans during the ice age
This is a very important and ground breaking paper by Mme Nicole Pailhaugue which summarises the fauna and seasons of occupation of the important site of La Grotte de la Vache, in the French Pyrenees. It deserves a much wider audience.

Entrance to La Grotte de La Vache
Photo: Don Hitchcock 2008
I have read many, many papers on archaeology. Most are, to be frank, somewhat pedestrian. Rarely do you find a paper which demonstrates not just the ability to cope with mind-numbing amounts of raw data, but a demonstration of the ability to put that data into a coherent whole, and then to mine the data for very important results.
This is such a paper.
I was thrilled as I read this paper to see what wonderful insights that Mme Pailhaugue has been able to read into what might otherwise have been a very ordinary set of results from a less skilled and professional researcher.
This is a superb piece of work.
The translation and presentation in web-ready form here took many hours of work, but it was worth every second. I hope that this web page will result in Mme Pailhaugue's results being available to a wider audience.
Quaternaire Volume 9, Numéro 4, 1998. pp. 385-400
The fauna found at la Grotte de la Vache, an important site of the Pyrenean Magdalenian, was completely studied for the first time. 29 species of mammals and 21 species of birds were recognised.
This rich fauna is a basic reference for the knowledge of the Late Glacial period in the Pyrenees, with a strong proportion of mountain species (Capra pyrenaica (Ibex), Rupicapra pyrenaica (Chamois), Lagopus mutus (Ptarmigan), Accipitridae (Hawks, Eagles, and Harriers), Pyrrhocorax graculus (Alpine Chough) and Montifringilla nivalis (White winged Snowfinch)).
Arctic species (Rangifer tarandus (Reindeer), Alopex lagopus (Arctic Fox ), Lupus timidus (Mountain Hare)) are much more frequent than temperate species (Cervus elaphus (Red Deer), Vulpes vulpes (Red Fox), Oryctolagus (European Rabbit), Perdix perdix (Grey Partridge), Coturnix coturnix (Common Quail), Garrulus glandarius (Eurasian Jay) or Picus viridus (Woodpecker)).
This fauna is essentially constituted of the remains of meals and results from non-selective hunting. Ibex and Ptarmigan provide the main part of the game. The proportion of males and females and the study of ages in the populations of Ibex, Chamois and Reindeer let us know the seasons when the site was occupied by the hunters and allow us to define the way they have used it.
Animals and Seasons of Occupation of the Salle Monique
from the Pyrenean Magdalenian of the Grotte de la Vache
Nicole Pailhaugue (1998)
Location and Archaeological Context
The Grotte de la Vache is on the left bank of the Vicdessos, two kilometres south of Tarascon-sur-Ariege where the Vicdessos joins the Ariège. Situated at the base of the limestone outcrops of the Massif de Génat (1077m) the cave is 100 metres above the current level of the river and provides an excellent vantage point: the view extends to Saut du Teil, a narrowing of the valley downstream, across the whole Niaux basin, and the surrounding limestone mountains which rise to around 1100 metres, and above the valley appear peaks of the Pyrenees chain, which in this sector has two peaks of 3000 metres.

The Magdalenian penetration of the Tarascon Basin occurred relatively late, well after 21 300 BP. The human settlement of this area in the Middle and especially the final Magdalenian is quite remarkable, since no fewer than 5 painted caves and dozens of sites were occupied at that time in a radius of about 7 km radius from Tarascon-sur-Ariege.
Description and History of the Excavations
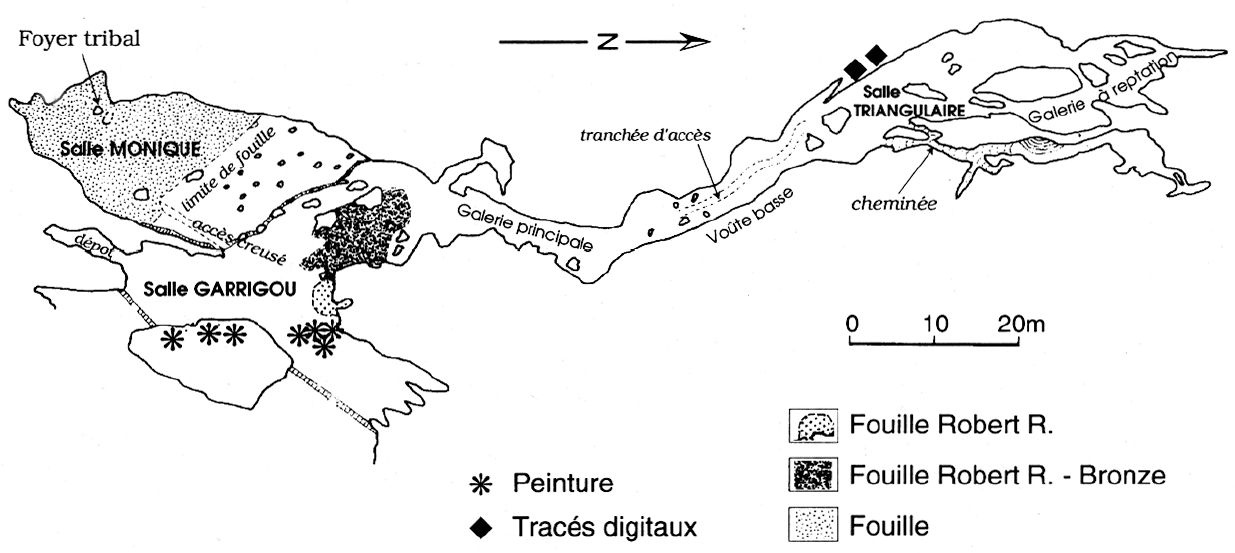
Fig. 1
Plan of la Grotte de la Vache, after the Spéléo-Club du Haut-Sabarthez
The cave is developed in the Urgo-Aptian limestone (the Urgo-Aptian is one of the stages of the Lower Cretaceous) following a south/north axis which corresponds to a sliding fault inside this formation (Fig. 1). Two openings, respectively oriented towards the east and southeast, light the Salle Garrigou. From there, a gallery extends northward sixty metres to reach the Salle Triangulaire beyond which further access is currently impossible. South of the Salle Garrigou there is an opening to the Salle Monique , also lit by natural light, but nevertheless easy to close off with a barrier of some kind if the occupants wanted better protection from the cold.
The cave has attracted the attention of prehistorians: F. Garrigou (1867), then F. Regnault (1872) have searched the entrance hall from the 1860s. Here F Garrigou demonstrated for the first time the presence of man from the Age of Reindeer in the Pyrenees.
From 1940, R. Robert resumed excavations in the Salle Garrigou. He then searched the Salle Monique during 1952-1964. From the faunal excavations of F. Garrigou, we have found to date only a few pieces that he added to the collections of the Prehistoric Museum of Ariège during its creation at the Château de Foix. The fauna of the excavation by R. Robert in the Salle Garrigou is only known by the list of taxa (Malvesin Fabre and Robert 1951). Our study (Pailhaugue, 1993, 1995) covers the material of the faunal excavations by R. Robert in the Salle Monique for which the only previously known data were due to the work of Dr. F.E. Koby (1957a, 1957b, 1959), who had had less than a small fraction of the materiel to work with.
Stratigraphy
In the Salle Monique R. Robert found the stratigraphy described by F. Garrigou (1867) and verified by him in the Salle Garrigou as follows, from top to bottom:
- A clay layer, sometimes absent, no more than 40 cm thick.
- A 'stalagmite' layer, of white calcium carbonate, of variable thickness.
- A layer of sand and pebbles
It is easy to see on the vertical faces of the excavation in the Salle Monique that sediments laid down at the same time are attributed to different layers. For example, layer IV at the bed of the large fireplace in the northwest is from the same lot of sediments as layer I at the back of the room. This is why we have not sought to highlight the stratigraphic division of the fauna.
Dating
Two carbon-14 dates on charcoal were performed by the laboratory of Groningue at the request of R. Robert:
Gr 2025: 12 540 ± 105 BP Layer II
Gr 2026: 12 850 ± 140 BP layer IV
A new dating performed at our request by the laboratory of Gif-sur-Yvette on a lot of bones confirmed the previous ones:
Gif. 7603: 12 800 ± 140 BP layer II
Recently, we have obtained dates produced by the particle accelerator laboratory in Gif-sur-Yvette:
Square 269, depth 109 cm: 13 490 ± 120 BP
Square 71, depth 59.5 cm: 13 770BP ± 140
Square 71 depth 115 cm: 13 650BP ± 130
These last three dates are perfectly compatible with each other. Located at the bottom of the Salle Monique, square 71 belongs to an area where the roof tends to collapse, dropping detritus, and where the number of remains is by far the highest in the entire gisement/site. This high number of remains occurred probably because it is an area bordering the living area that regularly accumulated extra material that was not thrown away outside the cave, because it constituted a reserve of raw materials in short supply, such as bones which might be able to be reworked into tools.
Human occupation of the deposit is contemporaneous with the Late glacial and presumably starting a little before. The only pollen analysis carried out to date (Leroi-Gourhan, 1967) is the anterior part of the deposit at the end of the Bölling and the start of the Dryas II. The lithic and bone industry contains thousands of pieces typical of the middle Pyrenean Magdalenian and the final Pyrenean, as well as a significant number of remains clearly attributable to the Azilian.
Fauna
La Salle Monique has delivered more than 142 000 animal remains identified to date (Table 1, Fig. 2-5), distributed among 29 taxa of mammals and 21 taxa of Birds.
The remains of fish and amphibians (such as frogs, toads, salamanders and newts) are not reviewed here.
Because of the diversity and the large number of data points, it is clear that this fauna is very representative of the extreme end of the Pleistocene in mountainous areas. Essentially, the remains have been the result of human action, as evidenced by the many examples of tools linked with the hunting and preparation of the animals. These tools are the most abundant types in the Tarascon Basin.
The birds, represented by a minimum of 4886 individuals (MNI frequency, MNI = Minimum Number of Individuals) far outweigh the numbers of mammals (MNI 2259, fig. 2). Among these, the Herbivores provided the largest share of the meat. (fig 3):
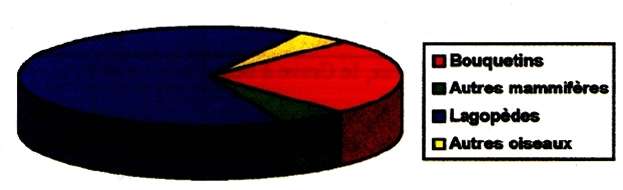
Fig. 2 - Pie Graph of major species found
Bouquetins - Ibex
Autres mammifères - Other mammals
Lagopèdes - Ptarmigan
Autres oiseaux - Other birds

Fig. 3 - Pie Graph of major mammals found
Bouquetins - Ibex
Rennes - Reindeer
Isards - Chamois
Renards - Foxes
Lièvres - Hares
Autres - Other mammals

Fig. 4 - Pie Graph of major birds found
Lagopèdes - Ptarmigan
Chocards - Alpine Chough
Perdrix - Partridges
Autres - Other birds

Fig. 5 - Pie Graph of some temperate and cold loving mammals apart from Ibex found
Renard commun - Red Fox
Cerf - Red Deer
Lapin - Rabbit
Renard polaire - Arctic Fox
Renne - Reindeer
Lièvre variable - Hare
Table 1 - Faunal list from La Salle Monique
| Species | Common Name | Number of remains identified | MNI = Minimum Number of Individuals |
|---|---|---|---|
| Mammals: | |||
| Carnivores: | |||
| Felidae: | |||
| Felis silvestris | Wildcat | 20 | 2 |
| Lynx sp. | Lynx | 31 | 2 |
| Panthera spelaea | Cave Lion | 1 | 1 |
| Canidae: | |||
| Canis lupus | Grey Wolf | 399 | 8 |
| Cuon sp. | Dhole, Wild Dog | 9 | 1 |
| Vulpes vulpes | Red Fox | min. 111 | |
| Alopex lagopus | Arctic Fox | min. 68 | |
| Indeterminate foxes | 1580 | 92 | |
| Ursidae: | Bears | min. 68 | |
| Mustelidae: | Weasels | ||
| Mustela erminea | Stoat or Ermine | 1 | 1 |
| Gulo gulo | Wolverine or Glutton | 12 | 2 |
| Artiodactyla: | |||
| Suidae: | Pigs | min. 68 | |
| Sus scrofa | Wild Boar | 33 | 2 |
| Cervidae: | deer | ||
| Cervus elaphus | Red Deer | 755 | 9 |
| Rangifer tarandus | Reindeer | 5821 | 147 |
| Capreolus capreolus | European Roe Deer | 14 | 2 |
| Bovidae: | Cattle, Bison, some antelopes, sheep, goats | ||
| Caprinae: | Sheep, Goats | ||
| Rupicapra pyrenaica | Chamois | 3307 | 85 |
| Capra pyrenaica | Ibex | 71451 | 1831 |
| Bovinae: | Cattle, bison, some antelopes | 206 | 6 |
| Perissodactyls: | Horses, Rhinoceroses | ||
| Equidae: | |||
| Equus caballus | Horse | 16 | 3 |
| Lagomorphes: | Hares, rabbits | ||
| Lepus timidus | Mountain Hare | 1058 | 42 |
| Oryctolagus cuniculus | European Rabbit | 63 | 5 |
| Rodentia: | Rodents | ||
| Citellus sp. | Ground squirrels | 2 | 1 |
| Marmota marmota | Alpine Marmot | 5 | 1 |
| Eliomys quercinus | Garden dormouse | 5 | 2 |
| Apodemus sylvaticus | Long-tailed field mouse | 4 | 2 |
| Arvicola sp. | Water vole | 5 | 3 |
| Microtus cf. arvalis | Common vole | 4 | 1 |
| Eulipotyphla: | Insectivores - Shrews, moles and hedgehogs | ||
| Talpa cf. europaea | European mole | 12 | 1 |
| Erinaceus cf. europaeus | European hedgehog | 18 | 4 |
| Total mammals: | 85029 | 2259 | |
| Species | Common Name | Number | MNI = Minimum Number of Individuals |
| Birds: | |||
| Anatidae: | |||
| Anas platyrhynchos | Mallard, or Wild Duck | 4 | 2 |
| Aythya fuligula | Tufted Duck | 3 | 1 |
| Mergus merganser | Common Merganser or Goosander | 18 | 7 |
| Anas clypeata | Northern Shoveler | 18 | 7 |
| Accipitridae: | |||
| Gypaetus barbatus | Lammergeier, or Bearded Vulture | 39 | 3 |
| Aquila chrysaetos | Golden Eagle | 11 | 2 |
| Tetraonidae: | |||
| Lagopus mutus, Lagopus lagopus | Rock Ptarmigan, Willow Ptarmigan | 52627 | 4566 |
| Phasianidae: | |||
| Perdix perdix | Grey Partridge | 540 | 91 |
| Coturnix coturnix | Common Quail | 4 | 1 |
| Columbidae: | Pigeons and Doves | 13 | 3 |
| Strigidae: | |||
| Nyctea scandiaca | Snowy Owl | 1 | 1 |
| Asio otus | Long-eared Owl | 2 | 1 |
| Turdidae: | Thrushes | 38 | 6 |
| Ploceidae: | |||
| Montifringilla nivalis | White-winged Snowfinch | 19 | 6 |
| Corvidae: | |||
| Garrulus glandarius | Eurasian Jay | 7 | 4 |
| Pica pica | European Magpie | 52 | 16 |
| Pyrrhocorax pyrrhocorax | Red billed Chough | 23 | 7 |
| Pyrrhocorax graculus | Yellow billed Chough or Alpine Chough | 1137 | 147 |
| Corvus corax | Common Raven | 23 | 7 |
| Total birds: | 54 724 | 4 886 | |
| Total Fauna: | 139 753 | 7 145 | |
The Pyrenean Ibex is by far the most hunted mammal, with 1831 individuals minimum (MNI obtained by adding the 398 D4 lower rights, and the 1433 M3 lower lefts).
- With a minimum of 147 individuals, the reindeer is 13 times less frequent than the Ibex in the table. This total of 147 individuals corresponds to:
- At least 37 reindeer aged more than 29 months represented by the upper and lower jaws carrying deciduous (milk) teeth.
- 113 M3 isolated inferior rights, which belong to three right mandibles of reindeer aged 15-29 months and from 110 subjects aged more than 29 months.
- 30 mandibles with deciduous teeth corresponding to subjects aged less than 30 months.
- 57 M3 lower rights, with 2 of the right mandibles of individuals aged 27-30 months and 55 isolated teeth from individuals aged at least 30 months.
Amongst the birds (Fig. 4), Ptarmigan, including Willow Ptarmigan and Rock Ptarmigan (Koby, 1957 b), are clearly dominant, with a minimum 4566 individuals, or 93.5% of the number of birds represented in the deposit. Then come the Chough (MNI 147) and the Grey Partridge (MNI 91).
The hunting list of the occupants of the Salle Monique fits perfectly in a mountain habitat: the Ibex population in Haute-Ariege was remarkably well developed, but not that of the Chamois, which probably had a real demographic expansion during the Holocene (the period which followed) when the improved, wetter weather conditions allowed the development of a forest canopy to which the animal appears quite strongly attracted. Other forms no less typical of mountains are present (Dejonghe, 1984): the Alpine Marmot, the Bearded Vulture, the Golden Eagle, the Vulture, the Chough, the Alpine Chough or the Snowfinch.
The clear superiority in numbers of the Ibex over the Chamois, the Deer over the Reindeer (MNI 9), Hare over the Rabbit (MNI 5) or again the presence of the Arctic Fox over the Red Fox (Fig. 5) infer a cold climate, even a frigid one, for at least part of the tenure by humans of the Salle Monique.
The animals of the forest such as Roe Deer, Boar, or Red Deer, are few in number and are concentrated mostly in the north-west of the excavation. Birds also, at least among the temperate species of Quail, Jay, or Woodpecker are very scarce.
The faunal composition varies across sectors of the excavation. We were able to highlight (Pailhaugue, 1993) areas of concentration for reindeer and also for Red Deer; also the north-east sector where there is a concentration of the remains of Red Deer supplied the majority of the remains of wild boar and all the remains of the Roe Deer. The artificial stratigraphy used by R. Robert does not permit the demonstration of any changes in the composition of the fauna during the period of occupation of the Salle Monique, nor allow us to place in time the appearance of one species or the disappearance of another.
Compared to the list of faunal remains of F. Garrigou (1867) numbering about 60 000 pieces and that of G. Malvesin-Fabre and R. Robert (1951) concerning the Salle Garrigou, the large number of faunal remains of the Salle Monique allows us to identify not just the same species but to enrich the list of taxa by a dozen species or subspecies not found in the other excavations.
Several carnivores rarely encountered in excavations have confirmed their presence in the late Glacial of the Pyrenees: a canine, the Dhole, which persisted much later in the Haute-Ariège, and among the big cats, the Lynx, the Wild Cat and the Cave Lion; a mustelid, the Wolverine, already reported in the neighbouring excavations of l'Herm (Ball, 1894), Les Trois Frères (Begouën and Koby, 1950) and at Labastide (Clot, 1982). A large Brown Bear was also present.
Bovines and Horses are very rare, these large herbivores do not frequent areas where the terrain does not suit them.
Rodents and Insectivores (Lavocat et al., 1966) are also scarce in the site, the presence of some of them may also result from an incursion of these cave dwelling animals into the Salle Monique when not occupied by humans.
Amongst birds, few Ducks, Geese and Swans came to the neighbourhood of the Vicdessos. Raptors, diurnal and nocturnal, are also rare. With the exception of Owls, they still exist in the Pyrenees, and the Grey Partridge now has an isolated population in Haute-Ariège.
Birds such as the Quail and the Woodpecker have been identified for the first time on a Pyrenean gisement (site) from the recent Würm (Clot and Morer-Chauauviré, 1986), and the Tufted Duck and the Jay, corresponding to a temperate climate, are very scarce. The Chocard, the Yellow Beaked Chough, by far the most common corvid, is better represented than the Crave, the Red Beaked Chough, and it seems to cope better with cold conditions.
Hunting Goats
The Pyrenean Ibex and the Chamois
The anatomical distribution of teeth (Table 2) allows us to recognise the presence of juveniles still having their deciduous (milk) teeth. Of the 1831 Ibex in the Salle Monique:
- 28% of the subjects still have the D4 lower right: so they were under the age of 28 months at the time of their death.
- 72% have the left lower M3 and were therefore older than 28 months at their death (Couturier, 1962).
In several parts measured, where the epiphyses (rounded end of a long bone) are either absent or isolated, or imperfectly welded to the shaft, or they come from young animals, we assigned a maximum age by referring to the data of R. Barone (1966) concerning the goat.
Table 2: La Salle Monique. Capra pyrnaica. Anatomical distribution of dental remains
| Upper Left Teeth | Upper Right Teeth | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dentition | D2 | D3 | D4 | P2 | P3 | P4 | M1 | M2 | M3 | D2 | D3 | D4 | P2 | P3 | P4 | M1 | M2 | M3 |
| Deciduous Teeth | 119 | 179 | 201 | 1 | 116 | 35 | 137 | 187 | 205 | 129 | 30 | |||||||
| Permanent Teeth | 277 | 398 | 452 | 431 | 344 | 175 | 250 | 393 | 430 | 440 | 338 | 148 | ||||||
| Single Teeth | 30 | 41 | 70 | 210 | 291 | 381 | 614 | 762 | 889 | 37 | 33 | 68 | 201 | 258 | 393 | 590 | 765 | 934 |
| Single Teeth | 20 | 188 | 1 | 14 | 179 | |||||||||||||
| Single Teeth | 60 | 62 | ||||||||||||||||
| Total | 149 | 220 | 271 | 487 | 690 | 833 | 1161 | 1141 | 1064 | 174 | 220 | 273 | 451 | 651 | 823 | 1159 | 1133 | 1082 |
| Total | 640 | 2030 | 3614 | 668 | 1939 | 3615 | ||||||||||||
| Total | 6284 | 6222 | ||||||||||||||||
| Uncategorised fragments | 350 | |||||||||||||||||
| Total upper teeth | 12856 | |||||||||||||||||
| Lower left teeth | ||||||||||||||||||
| Dentition | I1 | I2 | I3 | C | i1 | i2 | i3 | c | D2 | D3 | D4 | P2 | P3 | P4 | M1 | M2 | M3 | |
| Deciduous Teeth | 2 | 11 | 1 | 1 | 208 | 282 | 268 | 1 | 1 | 175 | 64 | |||||||
| Permanent Teeth | 5 | 20 | 41 | 34 | 307 | 814 | 901 | 860 | 767 | 480 | ||||||||
| Single Teeth | 2426 | 367 | 249 | 37 | 48 | 65 | 88 | 147 | 240 | 302 | 574 | 751 | 953 | |||||
| Single Teeth | 287 | 41 | ||||||||||||||||
| Single Teeth | 126 | |||||||||||||||||
| Total | 7 | 31 | 41 | 401 | 1 | 38 | 256 | 347 | 356 | 454 | 1055 | 1204 | 1609 | 1582 | 1433 | |||
| Total | 3193 | 1247 | 2713 | 4791 | ||||||||||||||
| Total lower left teeth | 11944 | |||||||||||||||||
| Lower right teeth | ||||||||||||||||||
| Dentition | I1 | I2 | I3 | C | i1 | i2 | i3 | c | D2 | D3 | D4 | P2 | P3 | P4 | M1 | M2 | M3 | |
| Deciduous Teeth | 7 | 14 | 6 | 1 | 3 | 6 | 10 | 223 | 318 | 303 | 4 | 2 | 193 | 75 | 1 | |||
| Permanent Teeth | 5 | 18 | 42 | 37 | 292 | 778 | 875 | 885 | 789 | 486 | ||||||||
| Single Teeth | 340 | 39 | 51 | 52 | 95 | 162 | 262 | 311 | 602 | 734 | 937 | |||||||
| Single Teeth | 365 | 157 | ||||||||||||||||
| Total | 12 | 32 | 48 | 377 | 1 | 3 | 6 | 49 | 274 | 370 | 398 | 454 | 1044 | 1188 | 1680 | 1598 | 1424 | |
| Total | 3033 | 1331 | 2687 | 4927 | ||||||||||||||
| Total lower right teeth | 11978 | |||||||||||||||||
| Uncategorised fragments | 83 | 223 | ||||||||||||||||
| Total lower teeth | 24228 | |||||||||||||||||
The dimensions of the distal region of the left humerus and imperfect welding of epiphyses to the diaphysis allows us to identify (Fig. 6):
- 44% as males.
- 56% as females and juveniles aged under one year of indeterminate sex.
- 43% as males.
- 57% as females and juveniles aged under three years, of indeterminate sex.
- 40% as males and 60% as females aged over 3 years.
From the right calcaneus we can recognise (Fig. 8):
- 29% as juveniles aged under 3 years of indeterminate sex.
- Among subjects older than 3 years, 41% as male and 59% as female - the proportion very close to that obtained for the anterior cannon bone.
The jaws of young goats older than 4 years, still possessing their deciduous teeth and all or part of their permanent teeth are numerous. With information available on the replacement of deciduous teeth by permanent teeth among modern populations, it was possible to divide the subjects under 4 years of Salle Monique into four age groups (Fig. 9 and 10):
- 202 kids ages 4 to 10 months.
- 124 male and female juveniles aged 17-22 months.
- 138 bucks and does aged 27-34 months.
- Some subjects aged 40-45 months, hardly recognisable because identification is based on the presence of the adult canine tooth, and mesial (towards the middle of the front of the jaw) portions of the mandible are rarely preserved
By the same method, and comparing the skulls of chamois from the current Réserve Nationale de Chasse d'Orlu killed at known ages, the chamois of the Salle Monique (Table No. 4) are divided into three age groups (Fig. 11):
- 16 kids aged from 6 to 12 months.
- 8 juvenile males and females aged 16 to 20 months.
- Six bucks and does aged 27-32 months.
- Abundant food
- Rocky slopes offering the potential for escape and areas of refuge in many directions
- Slopes too steep to hold much snow, something that the Chamois did not concern itself with
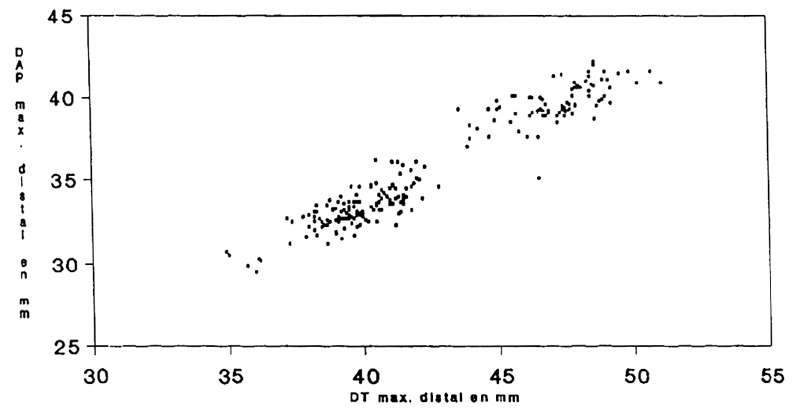
Fig. 6
La Salle Monique
Capra pyrenaica
Scatterplot of length versus width of the Ibex humerus bones found.
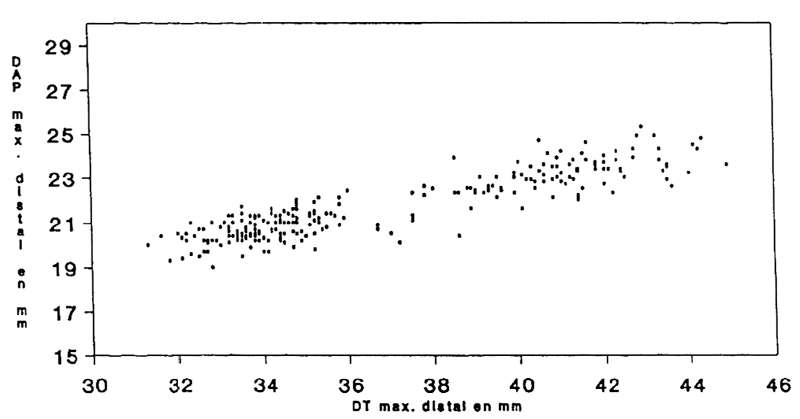
Fig. 7
La Salle Monique
Capra pyrenaica
Scatterplot of length versus width of the Ibex metacarpal bones found.
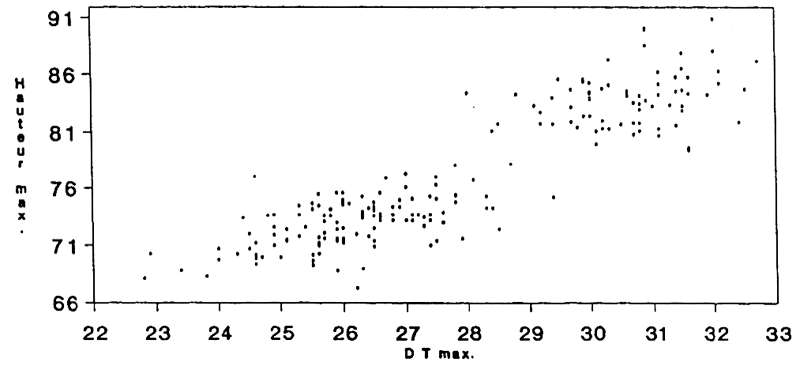
Fig. 8
La Salle Monique
Capra pyrenaica
Scatterplot of length versus width of the Ibex calcaneum bones found.
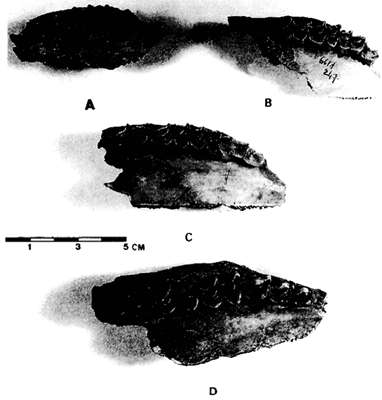
Fig. 9
La Salle Monique
Capra pyrenaica
Right Maxilaries
A and B: Animals between 4 and 10 months old, D2-M1.
C: Animal between 17 and 22 months old, D2-M2.
D: Animal between 31 and 34 months old, P2-M3.

Fig. 10
La Salle Monique
Capra pyrenaica
Left mandibles
A: Animal between 7 and 9 months old, D2-M1.
B: Animal between 18 and 22 months old, D2-M2.
C: Animal between 30 and 34 months old, P3-M3.

Fig. 11
La Salle Monique
Capra pyrenaica
A: Animal between 6 and 12 months old, right maxillary with D2-M1.
B: Animal between 28 and 30 months old, right maxillary wit P2-M3.
C: Animal between 6 and 12 months old, right mandible with D2-M1.
D: Animal between 16 and 20 months old, right mandible with D3-M2
E: Animal between 27 and 30 months old, right mandible with M2-M3 and left mandible with D4-M2.

Fig. 12
La Salle Monique
Rangifer tarandus
A: Animal between 3 and 5 months old, left maxillary with D2-M1.
B: Animal between 3 and 13 months old, right maxillary wit D2-M1.
C: Animal between 3 and 13 months old, right mandible with D2-M1.
D: Animal between 10 and 15 months old, left mandible with D3-M2.
E: Animal between 12 and 22 months old, left mandible with D3-M2.
F: Animal between 15 and 29 months old, right mandible with D3-M3.
Table 3: La Salle Monique. Capra pyrnaica. Anatomical distribution of bones
| Left | Right | Not assigned | Total | MNI = Minimum Number of Individuals | |||
|---|---|---|---|---|---|---|---|
| PF | DF | PF | DF | ||||
| Cranial fragments | 959 | 959 | |||||
| Bony horn sockets | 273 | 309 | 180 | 762 | 306 | ||
| Atlas (C1) and fragments | 90 | 90 | |||||
| Axis (C2) and fragments | 56 | 56 | |||||
| Scapula | 277 | 284 | 561 | 284 | |||
| Humerus | 6 | 694 | 9 | 645 | 1354 | 694 | |
| Radio-ulna | 356 | 128 | 362 | 121 | diaphysis 2 | 969 | 362 |
| Single ulna | 207 | 18 | 225 | 29 | 479 | 223 | |
| Lunate (carpal) | 379 | 369 | 748 | 379 | |||
| Supercarpal | 180 | 161 | 341 | 180 | |||
| Scaphoid (carpal) | 398 | 395 | 793 | 398 | |||
| Pyramidal (carpal) | 249 | 272 | 521 | 272 | |||
| Trapezoid (carpal) | 417 | 449 | 866 | 449 | |||
| Hamate (carpal) | 245 | 248 | 493 | 248 | |||
| Anterior cannon bone | 137 | 482 | 174 | 489 | E.D. 194 | 1476 | 489 |
| Metacarpals | 125 | 125 | |||||
| Coxa | 75 | 100 | 175 | 100 | |||
| Femur | 38 | 27 | 34 | 27 | 126 | 38 | |
| Patella | 194 | 260 | 454 | 260 | |||
| Tibia | 31 | 562 | 35 | 552 | diaphysis 1 | 1181 | 562 |
| Malleolus | 107 | 109 | 216 | 109 | |||
| Navicular bone | 379 | 328 | 707 | 379 | |||
| Talus | 792 | 803 | 1595 | 803 | |||
| Calcaneus | 488 | 438 | 926 | 488 | |||
| Calcaneus epiphysis not attached | 26 | 28 | 54 | 28 | |||
| Large cuneiform | 113 | 134 | 247 | 134 | |||
| Small cuneiform | 5 | 5 | 10 | 5 | |||
| Posterior cannon bone | 270 | 416 | 239 | 419 | E.D. 116 | 1460 | 419 |
| Small sesamoid bone | 22 | 22 | |||||
| Distal metapode fragments | 530 | 530 | |||||
| Large external sesamoid | 573 | 573 | |||||
| Large internal sesamoid | 538 | 538 | |||||
| 1st phalange and fragments | 6980 | 6980 | |||||
| 2nd phalange and fragments | 3201 | 3201 | |||||
| Small sesamoid | 247 | 247 | |||||
| 3rd phalange and fragments | 4532 | 4532 | |||||
| Total | 34367 | ||||||
PF: Proximal Fragments. DF: Distal fragments
Table 3: La Salle Monique. Capra pyrnaica. Anatomical distribution of bones
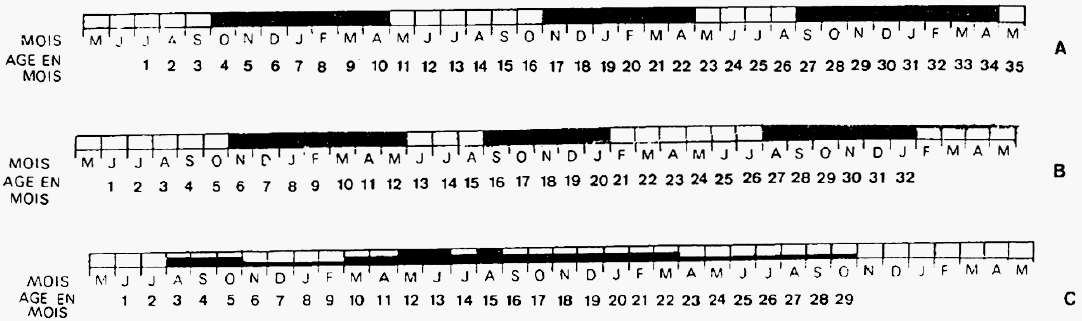
Fig. 13
La Salle Monique
Age groups and hunting seasons: A = Ibex, B = Chamois, C = Reindeer
Table 4: La Salle Monique. Rupicapra pyrenaica. Anatomic distribution of the dental remains.
| Upper Left Teeth | Upper Right Teeth | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dentition | D2 | D3 | D4 | P2 | P3 | P4 | M1 | M2 | M3 | D2 | D3 | D4 | P2 | P3 | P4 | M1 | M2 | M3 |
| Deciduous Teeth | 7 | 12 | 15 | 8 | 4 | 6 | 12 | 11 | 7 | |||||||||
| Permanent Teeth | 21 | 27 | 36 | 38 | 36 | 24 | 31 | 36 | 33 | 36 | 32 | 23 | ||||||
| Single teeth | 1 | 1 | 1 | 2 | 10 | 13 | 13 | 24 | 2 | 5 | 6 | 2 | 4 | 8 | 16 | 11 | 29 | |
| Single teeth | 3 | 9 | ||||||||||||||||
| Total | 36 | 260 | 42 | 277 | ||||||||||||||
| Uncategorised fragments | 3 | 12 | ||||||||||||||||
| Total Upper Teeth | 630 | |||||||||||||||||
| Lower left teeth | ||||||||||||||||||
| Dentition | I1 | I2 | I3 | C | i1 | i2 | i3 | c | D2 | D3 | D4 | P2 | P3 | P4 | M1 | M2 | M3 | |
| Deciduous Teeth | 10 | 21 | 22 | 12 | 4 | |||||||||||||
| Permanent Teeth | 29 | 49 | 49 | 43 | 38 | 20 | ||||||||||||
| Single Teeth | 19 | 9 | 3 | 2 | 2 | 1 | 2 | 1 | 2 | 4 | 7 | 20 | 32 | 34 | ||||
| Single Teeth | 7 | 1 | ||||||||||||||||
| Single Teeth | 14 | |||||||||||||||||
| Total | 38 | 5 | 12 | 22 | 22 | 31 | 53 | 56 | 75 | 74 | 54 | |||||||
| Total | 38 | 61 | 358 | |||||||||||||||
| Total lower left teeth | 457 | |||||||||||||||||
| Lower right teeth | ||||||||||||||||||
| Dentition | I1 | I2 | I3 | C | i1 | i2 | i3 | c | D2 | D3 | D4 | P2 | P3 | P4 | M1 | M2 | M3 | |
| Deciduous Teeth | 1 | 1 | 8 | 15 | 17 | 1 | 1 | 17 | 5 | |||||||||
| Permanent Teeth | 21 | 47 | 46 | 43 | 40 | 23 | ||||||||||||
| Single Teeth | 22 | 6 | 4 | 3 | 2 | 5 | 1 | 5 | 6 | 4 | 5 | 6 | 19 | 26 | 34 | |||
| Single Teeth | 7 | 2 | ||||||||||||||||
| Single Teeth | 22 | |||||||||||||||||
| Total | 2 | 1 | 1 | 13 | 15 | 23 | 25 | 53 | 53 | 79 | 71 | 57 | ||||||
| Total | 42 | 61 | 362 | |||||||||||||||
| Total lower right teeth | 465 | |||||||||||||||||
| Total uncategorised fragments | 5 | |||||||||||||||||
| Total lower teeth | 927 | |||||||||||||||||
Table 4: La Salle Monique. Rupicapra pyrenaica. Anatomic distribution of the dental remains.
Reindeer Hunting
Jaws of young reindeer can also define the age at death in months and thus the hunting seasons, but with less precision than those of Ibex and Chamois. (Delpech, 1983).
37 reindeer with their deciduous teeth were still under the age of 29 months at the time of their death. These young reindeer represent 25% of the population of reindeer in the Salle Monique, a percentage similar to those obtained for goats, probably also reflecting the natural composition according to the age of a population in which prehistoric indiscriminate hunting was practised.
They are divided into five age groups (Fig. 12):
- 3 reindeer aged 3 to 5 months
- 13 reindeer aged 3-13 months
- 7 reindeer aged 10-15 months
- 3 reindeer aged 12-22 months
- 11 reindeer aged 15-29 months
Examination of the frontal bone (the frontal bone of animals such as the reindeer is where horns or antlers are mounted. Male reindeer drop their antlers at the beginning of winter, and female reindeer retain their antlers until after they give birth in the spring) allowed the recognition of females culled during the moult in May-June, and males also killed during the moult that occurs during autumn. Most antlers are of males, which were necessarily collected by the hunters during the period of, or shortly after, moulting, the antlers quickly disappearing in the wild because some rodents are fond of them. These two observations confirm perfectly the presence of man in la Salle Monique in autumn and at the end of spring.
Assuming that the reindeers of the late Würm conducted micro-migration, probably at the regional level, all the reindeer from la Salle Monique could - it has not been demonstrated - have been hunted during their passage through the valley during the summer migration to the high pastures or during the descent of the herds returning to their winter territory in late summer/early autumn, when frost and snow make the high altitude feeding grounds inhospitable.
Table 5: La Salle Monique. Rupicapra pyrnaica. Anatomical distribution of bones
| Left | Right | Uncategorised | Total | MNI | |||
|---|---|---|---|---|---|---|---|
| PF | DF | PF | DF | ||||
| Cranial fragments | 86 | 86 | |||||
| Bony horn sockets | 13 | 19 | 10 | 42 | 19 | ||
| Atlas (C1) and fragments | 10 | 10 | |||||
| Axis (C2) and fragments | 15 | 15 | |||||
| Scapula | 11 | 17 | 28 | 17 | |||
| Humerus | 44 | 38 | 82 | 44 | |||
| Radio-ulna | 40 | 9 | 47 | 13 | 109 | 47 | |
| Single ulna | 29 | 24 | 3 | Diaphysis 1 | 57 | 29 | |
| Lunate (carpal) | 13 | 18 | 31 | 18 | |||
| Supercarpal | 4 | 6 | 10 | 6 | |||
| Scaphoid (carpal) | 19 | 10 | 29 | 19 | |||
| Pyramidal (carpal) | 2 | 11 | 13 | 11 | |||
| Trapezoid (carpal) | 15 | 15 | 30 | 15 | |||
| Hamate (carpal) | 4 | 7 | 11 | 7 | |||
| Anterior cannon bone | 9 | 28 | 5 | 41 | DF 21 | 104 | 41 |
| Lateral metacarpals | 12 | 12 | |||||
| Coxa | 11 | 7 | 18 | 11 | |||
| Femur | 1 | 3 | 1 | 4 | 9 | 4 | |
| Patella | 12 | 8 | 20 | 12 | |||
| Tibia | 3 | 26 | 5 | 22 | 56 | 26 | |
| Malleolus | 2 | 2 | 4 | 2 | |||
| Navicular bone | 22 | 21 | 43 | 22 | |||
| Talus | 29 | 24 | 53 | 29 | |||
| Calcaneus | 18 | 19 | 37 | 19 | |||
| Large cuneiform | 1 | 3 | 4 | 3 | |||
| Small cuneiform | 2 | 2 | |||||
| Posterior cannon bone | 10 | 19 | 16 | 28 | DF 17 | 90 | 28 |
| Distal fragments of the metapode | 32 | 32 | |||||
| Large external sesamoid | 7 | 7 | |||||
| Large internal sesamoid | 5 | 5 | |||||
| 1st phalange and fragments | 394 | 394 | |||||
| 2nd phalange and fragments | 160 | 160 | |||||
| Small sesamoid | 7 | 7 | |||||
| 3rd phalange and fragments | 140 | 140 | |||||
| Total | 1750 | ||||||
Table 5: La Salle Monique. Rupicapra pyrnaica. Anatomical distribution of bones
PF: Proximal Fragments. DF: Distal fragments
Table 6: La Salle Monique. Capra pyrnaica. Occurrence of bones with respect to Maximum Number of Individuals
| Left | Right | |||
|---|---|---|---|---|
| MNI | % MNI total | MNI | % MNI total | |
| Bony horn sockets | 306 | 16.7 | 275 | 15.0 |
| Atlas (C1) | 90 | 4.9 | ||
| Axis (C2) | 56 | 3.1 | ||
| Scapula | 275 | 15.0 | 284 | 15.5 |
| Humerus PF | 6 | 0.3 | 9 | 0.5 |
| Humerus DF | 694 | 37.9 | 645 | 35.2 |
| Radius PF | 356 | 19.4 | 362 | 19.8 |
| Radio-ulna DF | 128 | 7.0 | 119 | 6.5 |
| Single ulna PF | 206 | 11.3 | 223 | 12.2 |
| Single ulna DF | 18 | 1.0 | 29 | 1.6 |
| Lunate (carpal) | 379 | 20.7 | 369 | 20.2 |
| Supercarpal | 180 | 9.8 | 161 | 8.8 |
| Scaphoid (carpal) | 398 | 21.7 | 395 | 21.6 |
| Pyramidal (carpal) | 249 | 13.6 | 272 | 14.9 |
| Trapezoid (carpal) | 417 | 22.8 | 449 | 24.5 |
| Hamate (carpal) | 245 | 13.4 | 248 | 13.6 |
| Anterior cannon bone PF | 137 | 7.5 | 174 | 9.5 |
| Anterior cannon bone DF | 482 | 26.3 | 489 | 26.7 |
| Coxa | 75 | 4.1 | 100 | 5.5 |
| Femur PF | 38 | 2.1 | 34 | 1.9 |
| Femur DF | 27 | 1.5 | 27 | 1.5 |
| Patella | 194 | 10.6 | 260 | 14.2 |
| Tibia PF | 31 | 1.7 | 35 | 1.9 |
| Tibia DF | 562 | 30.7 | 489 | 26.7 |
| Malleolus | 107 | 5.8 | 109 | 6.0 |
| Navicular bone | 379 | 20.7 | 328 | 17.9 |
| Talus | 792 | 43.3 | 803 | 43.9 |
| Calcaneus | 488 | 26.7 | 438 | 23.9 |
| Posterior cannon bone PF | 270 | 14.7 | 239 | 13.1 |
| Posterior cannon bone DF | 416 | 22.7 | 419 | 22.9 |
| 1st phalange and fragments | about 27.0 | |||
| 2nd phalange and fragments | about 19.3 | |||
| 3rd phalange and fragments | about 30.9 | |||
Table 6: La Salle Monique. Capra pyrnaica. Occurrence of bones with respect to Maximum Number of Individuals
PF: Proximal Fragments. DF: Distal fragments
Table 7: La Salle Monique. Rupicapra pyrnaica. Anatomical distribution of bones vs MNI
|
Left MNI | Left % MNI Total | Right MNI | Right % MNI Total | |
|---|---|---|---|---|
| Bony horn sockets | 13 | 15 | 19 | 22 |
| Atlas (C1) | n = 10 % = 12 | |||
| Axis (C2) | n = 15 % = 18 | |||
| Scapula | 11 | 13 | 17 | 20 |
| Humerus Proximal Fragments | 44 | 52 | 38 | 45 |
| Radio-ulna Proximal Fragments | 40 | 47 | 47 | 55 |
| Single ulna | 29 | 34 | 24 | 28 |
| Lunate (carpal) | 13 | 15 | 18 | 21 |
| Supercarpal | 4 | 5 | 6 | 7 |
| Scaphoid (carpal) | 19 | 22 | 10 | 12 |
| Pyramidal (carpal) | 2 | 2 | 11 | 13 |
| Trapezoid (carpal) | 15 | 18 | 15 | 18 |
| Hamate (carpal) | 4 | 5 | 7 | 8 |
| Anterior cannon bone PF | 9 | 11 | 5 | 6 |
| Anterior cannon bone DF | 3 | 4 | 4 | 5 |
| Coxa | 11 | 13 | 7 | 8 |
| Femur PF | 1 | 1 | 1 | 1 |
| Femur DF | 3 | 4 | 5 | 6 |
| Patella | 12 | 14 | 8 | 9 |
| Tibia PF | 3 | 4 | 5 | 6 |
| Tibia DF | 26 | 31 | 22 | 26 |
| Malleolus | 2 | 2 | 2 | 2 |
| Navicular bone | 22 | 26 | 21 | 25 |
| Talus | 29 | 34 | 24 | 28 |
| Calcaneus | 18 | 21 | 19 | 22 |
| Large cuneiform | 1 | 1 | 3 | 4 |
| Posterior cannon bone PF | 10 | 12 | 16 | 19 |
| Posterior cannon bone DF | 19 | 22 | 28 | 33 |
| 1st phalange and fragments | n = 236 % = 36.42 | |||
| 2nd phalange and fragments | n = 128 % = 19.75 | |||
| 3rd phalange and fragments | n = 140 % = 21.61 | |||
PF: Proximal Fragments. DF: Distal fragments
Table 7: La Salle Monique. Rupicapra pyrnaica. Anatomical distribution of bones vs MNI
Animal Preparation Methods
The anatomical distribution of the remains of the Pyrenean Ibex (Table 2 and 3), Chamois (Table 4 and 5) and Reindeer shows that all skeletal elements are present in the deposit and the animals were most often if not always carried entire to the site to be skinned and butchered. Perhaps however they were already gutted because only one bovid bone (goats are part of the bovid group) from the foetal stage was found. We have now shown that most goats were slaughtered in winter or spring, periods when females are pregnant.
The representativeness of the main sets of bones compared to MNI often shows a remarkable coincidence of paired bones for the anatomical element between the right and its left counterpart (Table 6 and 7). The element of the post-cranial skeleton that is best represented is the astragalus or talus bone (43% of MNI) for the Ibex and the proximal radio-ulnar for the Chamois (55% of MNI), an occurrence of less than half that of the teeth, partly due to better natural conservation of the teeth, but also because of the processing of the bones by the hunters.
In early research at La Grotte de La Vache, Garrigou (1867) and Regnault (1872) found that the fragmentation of the bones could best be explained not by natural causes, but by human intervention. They relied for that conclusion on the data collected for the amount of fragmentation demonstrated for fossilised human bones in the Pyrenees. Koby also reported the presence of incisions due to the action of a bone tool, and systematic fragmentation of the long bones of ruminants.
Mandibles and long bones were fractured in a completely routine manner to extract the marrow. This practice also involved getting the marrow from most of the first phalanxes, which contain very little marrow (only 431 are complete out of over 3000 represented in the pool) and about half of the second phalanges, a short bone which contains much less marrow. Among the limb bones (phalanges excluded) of 1831 Ibex and the 85 chamois in the deposit, only two radio-ulnar bones and three cannon bones were found complete or nearly so. We do not mean to suggest, like Koby (1957a), that the prehistoric occupants of la Salle Monique suffered from "chronic hunger" or famine. The meat diet does not appear to have failed, given the wealth of the kill, but the marrow would have, in the form of animal fat, had many uses in addition to nutrition.
The tool marks on the skeletons of the Caprinae show that the stripping was carried out in order to keep the skin whole and entire by cutting at the carpus or metacarpus of the foreleg, and the tarsal or metatarsal of the hind leg. The skin was also cut around the base of the horns and at the upper lips, while the head was perhaps already separated from the trunk behind the occipital condyles. The limbs were separated from the carcass and cut at each joint. The dislocation of the spine and the limbs was very thorough, as far as the area of the autopode, an area rich in ligaments and tendons, which, after preparation, may provide thread for sewing. Meat and fat were then eaten, or prepared for deferred consumption, possibly by smoking or drying.
Conclusions
The Ibex has provided the staple diet for the prehistoric people of la Salle Monique. Its abundance has probably conditioned the organisation of a timetable for hunting, and the presence of man in the cave is demonstrated during a part of autumn, during winter and during early spring.
The chamois is much rarer than the ibex in the environment of the hunters from La Grotte de la Vache. It seems to have been hunted only occasionally during the same seasons as the ibex. This is hardly surprising, since both species can occupy the same wintering grounds and their seasonal behaviour is essentially the same if one refers to observations of known current populations.
The season of occupation at the Grotte de La Vache corresponds to the best times for hunting in mountain areas. Hunters in La Salle Monique enjoyed an excellent shelter during the cold season, during which time they were sure to get game in quite sufficient quantity for their needs in the immediate vicinity of the site. They knew very well the ethology of the animals that formed the basis of their food supply, and which supplied many of their raw materials. Their hunting calendar illustrates this knowledge.
During the summer, large groups of animals dispersed, the males first, then the does, going to the high altitude pastures. The hunt in summer would then have been very long and arduous, and the success of their hunting spears and atlatl thrown darts, very random. The lengthening of the days during the summer period was conducive to regional scale displacements allowing them to vary their food supplies and procure raw materials which were locally rare, particularly flint, or non-existent locally, such as the Atlantic and Mediterranean shells used to decorate clothing.
It is, however, certain that the prehistoric hunters were back in time for the start of the new season in time to hunt the reindeer returning to their wintering grounds. It is during the autumn migration that male reindeer shed their antlers, which were then picked up to provide almost all of the essential raw material for the renewal of spearheads and harpoons. This was also the period of salmon fishing on the spawning grounds of the Upper Ariège in autumn.
La Grotte de la Vache has yielded numerous objects made of bone or Cervidae antler, decorated with engraved or carved animals, and if the ibex (Fig. 1) is frequently represented, it does not appear that the chamois is featured on these items. The prehistoric representation of chamois is very rare, from memory consisting of a complete chamois and chamois head carved on the walls of the Grotte du Ker Massat, and engraved bones from the Grotte de Labastide. Deer, bison, aurochs, frequently appear in the portable art of the Grotte de la Vache, which also offers us the images of animals very rarely represented elsewhere, such as wolves, chum salmon, panthers, bears and even the head of a bird of indeterminate species carved on reindeer antler.
The Magdalenians of the Grotte de La Vache were assured of obtaining game in quite sufficient quantities in the immediate vicinity of the site, without major difficulties we think, probably by hunting parties in the case of herbivores and by trapping for small animals, particularly grouse and animals hunted for their fur. They benefited from the excellent shelter provided by the Salles Garrigou and Monique, which were illuminated by sunlight during the day but still easy to close off when better protection against the cold was necessary. These hunters appear to us to be a group of true mountaineers who were familiar with and used effectively the abundant resources of a mountain environment which is often regarded as difficult today.
Perhaps they were the first to learn to adapt to the environment of a high valley in Ariège which they occupied during at least two-thirds of the year. Witness the remains of all kinds found by the thousands, even tens of thousands, of wildlife in La Salle Monique, and more than two hundred art objects, which are all artistic masterpieces.
Bibliography
- Altuna, J., 1978: Dimorphisme sexuel dans le squelette postcéphalique de Capra pyrenaica pendant le Würm final, Munibe, 30, 4, 201-214
- Barone, R., 1966: Anatomie comparée des Mammifères domestiques, Ostéologie, I, 761 p.
- Begouen, H., Koby, F., 1950: Le crâne de Glouton de la caverne des Trois Frères (Ariège), Bull. S.P.A.P., V, 49-68
- Boule, M., 1894: Note sur des restes de Glouton et de Lion fossiles de la caverne de l'Herm (Ariège), Anthropologie, V, 10-14
- Clot, A., 1982: Le Glouton (Gulo gulo L., Mustelidae, Carnivora) de Labastide (Hautes-Pyrénées), Bull. Sté Hist. Nat. Toulouse, 118, 101-109
- Clot, A., Mourer-Chauvre, C., 1986: Inventaire sytématique des Oiseaux quaternaires des Pyrénées françaises, Munibe, 38, 171 - 184
- Couturier, M., 1938: Le Chamois, B. Arthaud Ed., Grenoble, 855 p.
- Couturier, M., 1962: Le Bouquetin des Alpes, Imprimerie Allier, Grenoble, 1564 p
- Dejonghe, J., 1984: Les Oiseaux de montagne, Ed. du point vétérinaire, Maisons-Alfort, 310 p.
- Delpech, F., Le Gall, O., 1983: La Faune magdalénienne de la grotte des Eglises (Ussat, Ariège), Bull. S.P.A.P, XXXVIII, 91 - 118
- Delpech, F., 1983: Au Sujet des migrations de rennes, Les faunes du paléolithique supérieur dans le Sud-Ouest de la France, Cahiers du Quaternaire, No 6, 164-174
- Delpech, F., 1983: Au Sujet des migrations de rennes, Les faunes du paléolithique supérieur dans le Sud-Ouest de la France, Cahiers du Quaternaire, No 6, 164-174
- Garrigou, F. , 1867: L'Age du Renne dans la grotte de La Vache, Bull. de la Sté d'Hist. Nat. de Toulouse, 11 p.
- Koby, F., 1957 a: La Faunule aviaire de la grotte de La Vache, Bull. S.P.A.P., XII, 79-96
- Koby, F., 1957 b: Les Lagopèdes de la station magdalénienne de La Vache dans les Pyrénées, Eglogae geologicae Helvetiae, 50, No2, 565-568
- Koby, F., 1959: Les Renards magdaléniens de La Vache avec remarques sur le diagnostic dentaire différentiel des genres Vulpes et Leucocyon, Bull. S.P.A.P., XIV, 26-34
- Lavocat, R. et al., 1966: Faunes et flores préhistoriques de l'Europe occidentale, Atlas de préhistoire, Boubée ed., III, 478 p.
- Leroi-Gourhan, A., 1967: Pollens et datations de la grotte de la Vache (Ariège) Bull. S.P.A.P., XXII, 115-127
- Malvesin-Fabre, G., Robert, R., 1951: Engins de chasse et de pêche du magdalénien de la grotte de La Vache (Ariège), Bull. S.P.A.P., VI, 13-30
- Pailhaugue, N., 1995: La faune de la Salle Monique, Grotte de La Vache (Alliat, Ariège), Bull. Soc. Préhist. Ariège-Pyrénées, L, 225
- Regnault, F., 1872: Sur les fouilles pratiques dans la grotte de La Vache, près de Tarascon (Ariège), Bull. Sté d'Anthropologie de Paris, 7, 202-204
- Taillefer, F., 1985: Idées actuelles sur les glaciations dans les Pyrénées de l'Ariège, Revue géographique des Pyrénées et du Sud-Ouest, 56, fasc. 3, 3232-338