Back to Don's Maps
Back to the Iron Gates - Location of the Sharamudoi
Vlasac
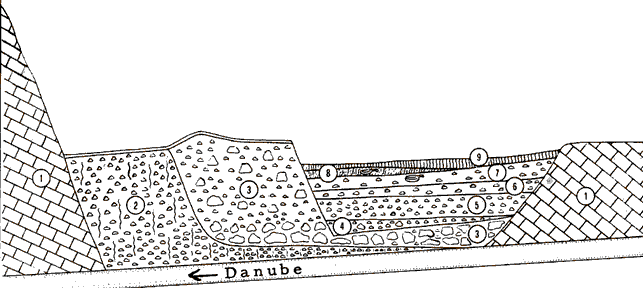
The habitations at Vlasac were semi dugouts with the floors made of limestone plaster. Rectangular hearths were constructed along the central axis of the habitations.
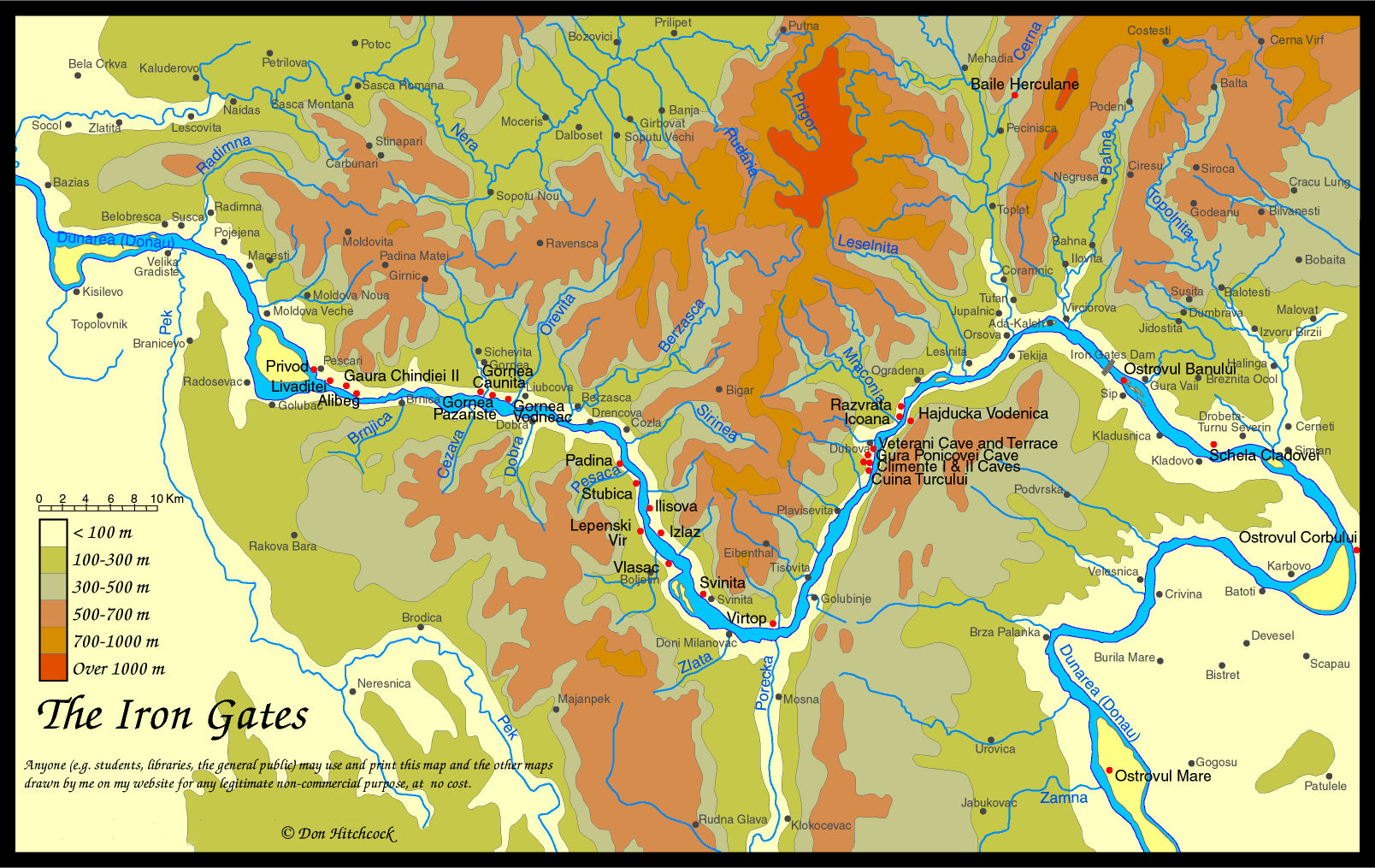
Map of the Iron Gates region, with the position of Vlasac marked.
Photo: Don Hitchcock 2014
Stratigraphy
Nine strata, including the basement rock, are described for the. Vlasac terrace, beginning with the oldest, number 1. The profile above provides a vertical profile of the relations between these strata (Markovic-'Marjanovic, 1978; SrejoviC' and Letica, 1978).
Stratum I
Stratum I consists of steeply tilted limestone bedrock which makes up the Boljetin Hill which lies directly behind the site. This stratum is exposed in some places above the site, and is the 'source of scree and boulders which occur in later strata. A meander, 232 meters across, was cut into this stratum by the Danube.
Stratum II
During the early Riss glaciation the meander cut into Stratum I began to fill with scree and loess and formed Stratum II. This Stratum is bright yellow in color. During the Wurm Interstadial, the water level in the Danube rose, and the river cut into Stratum II, forming a new depression 20 meters deep and over 100 meters wide. What remains of Stratum II varies in thickness from .5 to 15 meters and is 100 meters long by 10 meters deep.
Stratum III
In the early Wurm the depression cut into Stratum I , I began to fill with large polygonal blocks of Jurassic limestone and smaller scree, all with sharp edges, in a matrix of loess and formed Stratum III. Stratum III is mottled yellow in color. During the next interstadial, the Danube once again rose and cut another depression, this time into Stratum III. This depression is about 100 meters long. What remains of Stratum III varies in thickness from 1-15 meters and is 182 meters long by 20 meters deep.
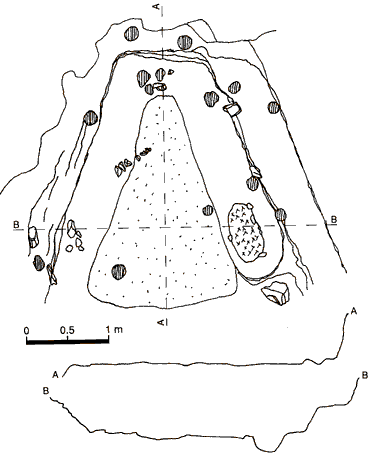
Stratum IV
Stratum IV is the first stratum containing cultural remains, those assigned to Vlasac Ia. Trapezoidal house floors with a red limestone pavement, excavated fireplaces, bone and stone are mixed with scree. The stratum has the mottled yellow color of the scree. Stratum IV is only found in the lowest depressions of the cut-out base on which it lies. It is not found in the middle area of the terrace, and occurs rarely in the SW periphery. It varies in thickness from .0 to .70 meters.
Stratum V
Stratum V contains the cultural remains of Vlasac Ib. These include red trapezoidal house floors, stone hearths, and a great amount of fish bone. These cultural remains are mixed with scree and hardened loess in a brown matrix. It is found in the same areas of the site where Stratum IV is found, and varies in thickness from .10 to 1.80 meters.
Stratum VI
Stratum VI contains the cultural remains of Vlasac II, which include circular stone rings (tent rings?), houses, hearths made of plaque-like stones, stone and bone. It is a dark brown, rich humus layer filled with large blocks and scree and varying,from .10 to 1.40 meters in thickness. It is much more extensive than Strata IV and V. occupying part of the middle area of the terrace, and more of the periphery, and is the overall thickest culturebearing level.
Stratum VII
Stratum VII occurred in every square which was excavated, and is thus the most extensive culture-bearing level. It is thickest in the western and central sectors. The cultural remains include stone rings, hearths made of crude blocks, bones and stones, belonging to Vlasac III. This level is a dark brown soil containing occasional large limestone blocks. In some places it has been damaged or destroyed by later (Neolithic) inhabitants. It varies in thickness from .20 to 1.00 meters.
Stratum VIII
Stratum VIII is a dark yellowish level with scattered lenses of Neolithic and later cultures. It varies in thickness from .15 to .80 meters.
Stratum IX
Stratum IX is a dark red or brown top soil containing occasional pieces of scree. Its thickness varies from .20 to .30 meters.
The following text comes from B. Prinz, 'Mesolithic Adaptations on the Lower Danube':
Eighty-four graves at Vlasac produced skeletal material from 117 individuals. Scattered finds of another 65 individuals came from all over the occupied area (Nemeskeri, 1978, pp.97-98). The general character of these remains, representing a tall (females 163.32, and males 170.53 cm), robust, Cro-Magnon-type population, with a tendency toward some gracilization as the, Mesolithic period drew to a close, (Nemeskeri and Szathmary, 1978, pp.178, 179) is consonant with the reports of the 37 individuals recovered from 30 graves at Padina (Zivanovic, S., 1974, p.153), and the 85 skeletons from Lepenski Vir (Nemeskeri, 1972, p.200). Gracilization, particularly of males, is a general trend throughout Europe in the late Upper Paleolithic and Mesolithic (Frayer, 1978).
An odd feature of the Vlasac skeletal population is the condition of the clavicles. Clavicles are typically robust for both sexes. 'It is peculiar that the robustness of the left side is more marked in both males and females.' (Nemeskeri and Szathmary, 1978, 1972). What task necessitates the heavier use of the left over the right arm for both left and right-handed people? Probably the task or tasks involved were the same for both sexes, and regularly and frequently performed ones. Fishing, typically done by both sexes, and certainly a regular and frequent activity at Vlasac, comes to mind, though there probably were other 'options' to account for this condition.
Lengyel (1978, p. 275) has analyzed the Vlasac bone for ABO blood groups, and concluded that males and females, on the basis of the ABO system, appear to be from different populations. The females at Vlasac fit right in with the Lepenski Vir Mesolithic population, whereas the percentages of AB for the males are quite distinct. Perhaps males with blood type AB were buried outside the settlement. Schwartz (personal communication) has pointed out that collapsing the 1,000-year time span into a single sample completely invalidates all conclusions about evolutionary changes in genetic frequencies or the existence of more than one gene pool.
Evidence for Population Replacement
Human skeletal material offers direct evidence relevant to the question of population replacement or continuity from the Mesolithic to Neolithic periods.
Only Lepenski Vir provided early Neolithic skeletal material which could be compared with the abundant earlier data. Starcevo skeletons are 'moderately tall' and tend to be more gracile than skeletons from the preceding period, although gracile and robust variants are described for this period (Nemeskeri, 1972, p.201, p.202). No argument is made or suggested for evidence of population displacement or admixture of a formerly distinct gene pool.
Diet and Nutrition
Evidence discussed here for the use of vegetable food resources at Vlasac is relevant to an understanding of the nutritional status of the population. The only major disease in the skeletal population is rickets, or osteomalacia. This is interpreted as evidence of D deficiency, which is incorrect. The predominance of fish in the diet rules out this possibility. Rickets in modern populations is usually associated with D deficiency because D is naturally available in few places (fish and sunshine) whereas calcium is everywhere (dairy products, green vegetables, nuts, grains and legumes). Most of the dietary calcium in our society comes from dairy products, which were unavailable in the Mesolithic. At Vlasac nuts and seeds would have been available in late summer and fall, and could have been stored through the winter.
Legumes and green leafy vegetables could not have been stored, and would have been available in late spring and summer. The Chenopodiaceae, which are important in all but one sample, are an exception to the rule that green leafy vegetables provide good sources of calcium. Chenopodiaceae, members of the goosefoot family like spinach, contain oxalic acid, which binds calcium and makes it unavailable for absorption. Probably the Vlasac populations suffered seasonal calcium deficiencies in the late winter and early spring when stored nuts had run out and fresh greens had not yet become available. This deficiency would have been aggravated by heavier reliance on chenopodia. In fact, the extensive evidence for calcium deficiency at Vlasac strengthens the hypothesis that chenopodia were probably quite important.
Prehistoric Fauna from Vlasac
From: Prinz, B., 1987 'Mesolithic Adaptations on the Lower Danube'
Some idea of the labour involved in archaeology may be gleaned from the fact that nearly 30 000 animal bones were dug out, sorted and classified on just this one site. The work involved is mind boggling.
No = number of bones
MNI = Minimum Number of Individuals
MNI represents an attempt to work out how many different animals there are in a deposit. If there are three skulls, for example, then there are obviously three separate animals. However if for example a left thigh bone and a right thigh bone are found in distinct layers, it is also reasonable to allocate them to different animals, and count them as two individuals. This is important when trying to work out the role of various animals in the economy of the site. The MNI is artificially reduced if careful excavation procedures are not followed. In addition, smaller animals are under represented if the deposits are not sieved.
| Taxon | Common Name | No. | % | MNI | |
| Domestic | |||||
| Canis familiaris | dog | 1914 | 6.6% | 160 | |
| Wild | |||||
| Ungulates | |||||
| Bos primigenius | aurochs | 54 | 0.2% | 4/5 | |
| Rupicapra rupicapra | chamois | 22 | 0.1% | 4 | |
| Cervus elaphus | red deer | 6732 | 23.1% | 90 | |
| Capreolus capreolus | roe deer | 510 | 1.8% | 19 | |
| Sus scrofa | wild pig | 1185 | 4.1% | 28 | |
| Carnivores | |||||
| Felis silvestris | wild cat | 45 | 0.2% | 7/8 | |
| Lynx lynx | lynx | 5 | 0.02% | 2 | |
| Mustelidae | weasel | 8 | 0.03% | ||
| Martes martes | pine marten | 248 | 0.9% | 38 | |
| Meles meles | badger | 58 | 0.2% | 8 | |
| Ursus arctos | brown bear | 168 | 0.6% | 10 | |
| Canis lupus | wolf | 103 | 0.4% | 10 | |
| Vulpes vulpes | red fox | 30 | 0.1% | 6 | |
| Small carnivore | 13 | 0.04% | |||
| Rodents and Insectivores | |||||
| Castor fiber | beaver | 71 | 0.2% | 9 | |
| Sciuris vulgaris | squirrel | 5 | 0.02% | 3 | |
| Lepus europaeus | brown hare | 22 | 0.1% | 4 | |
| Erinaceus europaeus | hedgehog | 1 | 0.003% | 1 | |
| Birds | |||||
| Pelecans cf. onocratalus | white pelican | 3 | 0.01% | ||
| Phalacrocorax carbo | cormorant | 3 | 0.01% | ||
| Egreta alba | great white egret | 2 | 0.01% | ||
| Anas platyryncho | mallard | 2 | 0.01% | ||
| Anas creca | teal | 2 | 0.01% | ||
| Aythya nyroca | ferrugionous duck | 1 | 0.003% | ||
| Aquila heliaca | imperial eagle | 6 | 0.02% | ||
| Milvus migrans | black kite | 5 | 0.02% | ||
| Haliaetos albicilla | white tailed eagle | 27 | 0.1% | ||
| Strix aluco | tawny owl | 2 | 0.01% | ||
| Carullus glanduarius | jay | 1 | 0.003% | ||
| Pica pica | magpie | 4 | 0.01% | ||
| Corvus corax | raven | 4 | 0.01% | ||
| Unidentified birds | 87 | 0.3% | |||
| Reptile | |||||
| Emys orbicularis | pond tortoise (some showed burning and butchering marks) | 317 | 1.1% | ||
| Fish | |||||
| Cyprinus carpio | carp | 1552 | 5.3% | ||
| Cyprinidae | 5230 | 18.0 | |||
| esox lucius | pike | 11 | 0.04% | ||
| Siluris glanis | catfish | 2283 | 7.8% | ||
| Other fishes | unidentified | 8372 | 28.8% | ||
| ------- | |||||
| 29108 |